|
"Помехоустойчивость" вирусов
В. И. АГОЛ
Московский государственный университет им. М.В. Ломоносова
Вирусы могут проявлять поразительную устойчивость к разнообразным
неблагоприятным внешним факторам. Более того, во многих случаях они легко приспосабливаются
к новым условиям, например вредоносным химическим соединениям, в том числе даже
таким, с какими им ранее не приходилось встречаться (например, к новым лекарствам).
Практическое значение живучести вирусов очевидно. Не менее интересны фундаментальные
закономерности, лежащие в ее основе. В статье использованы примеры, почерпнутые
преимущественно из данных изучения двух групп вирусов: пикорнавирусов и аденовирусов.
Пикорнавирусы - это мелкие вирусы животных, у которых строго
упорядоченная (икосаэдрическая) белковая оболочка обволакивает однонитевой РНК
- геном длиной 7,5-8 тыс. нуклеотидов (например, возбудители полиомиелита,
гепатита А, ящура). Аденовирусы,
вызывающие, в частности, острые респираторные заболевания человека (ОРВИ,
ОРЗ), крупнее, также имеют икосаэдрическую оболочку, и их геном
представлен двунитевой молекулой ДНК, в которой около 36 тыс.
пар нуклеотидов.
ВИДОВАЯ УСТОЙЧИВОСТЬ
Структура конкретного вируса хорошо приспособлена к условиям, существующим в
его экологической нише. Например, некоторые пикорнавирусы и
аденовирусы размножаются преимущественно в желудочно-кишечном тракте. Соответственно
они не обращают внимания на кислый рН желудочного сока, пищеварительные
протеолитические ферменты, детергенты желчи
и другие агрессивные факторы. Такая устойчивость действительно поразительна:
ведь, чтобы вызвать инфекционный процесс в чувствительной клетке, эти вирусы
должны освободить заключенную в белковой оболочке нуклеиновую кислоту при температуре
тела, нейтральном рН и других мягких физиологических условиях. Другими словами,
эти вирусы в зависимости от обстоятельств либо исключительно стабильны, либо
чрезвычайно лабильны. Это парадоксальное сочетание свойств обусловлено биологически
целесообразным устройством белковой оболочки (капсида) вирусов.
Хотя капсиды всех пикорнавирусов построены по общему плану, кислотоустойчивость
характерна именно для тех представителей этой группы, которые размножаются в
кишечнике. Другие пикорнавирусы (например, риновирусы также возбудители ОРВИ,
паразитирующие в клетках слизистой верхних дыхательных путей), напротив, легко
разрушаются при небольшом сдвиге рН в кислую сторону. Таким образом, детали
вирусной архитектуры очень важны.
Яркий пример биологической "предусмотрительности" дает механизм репликации аденовирусной
ДНК. Дело в том, что в молекуле ДНК есть особо уязвимые участки - ее концы.
Существует класс клеточных ферментов (так называемые экзонуклеазы), разрушающих
именно концы молекул нуклеиновых кислот. Однако у аденовирусов поврежденные
концы ДНК могут регенерировать. В линейных цепочках нуклеиновых кислот различают
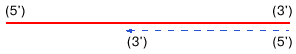
5'- и 3'-концы. Биосинтез новой цепи всегда идет в 5' 3'-направлении. Поскольку
в двунитевой структуре цепи обычно антипараллельны (имеют противоположную направленность),
матричная цепь всегда считывается в 3'  5'-направлении
(рис. 1). У аденовирусов есть по крайней мере два разных способа восстановить
утраченную концевую последовательность нуклеотидов. Один из них заключается
в следующем. Утеря конца двунитевой молекулы означает, что на одной из нитей
укорочен 5'-конец, а на другой - 3'-конец. При репликации генома этого вируса
происходит образование промежуточных однонитевых молекул ДНК. Нуклеотидная последовательность
обеих полноразмерных однонитевых молекул такова, что их концы взаимно комплементарны
и поэтому способны взаимодействовать друг с другом, образуя структуру, называемую
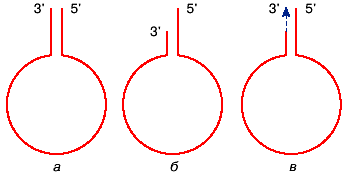
"ручкой сковородки" (рис. 2, а). Регенерация утерянного по какой-либо причине
3'-конца цепи (рис. 2, б ) большой проблемы для вируса не представляет, поскольку
структура полноразмерной ДНК может быть восстановлена в результате копирования
5'-конца этой же молекулы (рис. 2, в). 5'-направлении
(рис. 1). У аденовирусов есть по крайней мере два разных способа восстановить
утраченную концевую последовательность нуклеотидов. Один из них заключается
в следующем. Утеря конца двунитевой молекулы означает, что на одной из нитей
укорочен 5'-конец, а на другой - 3'-конец. При репликации генома этого вируса
происходит образование промежуточных однонитевых молекул ДНК. Нуклеотидная последовательность
обеих полноразмерных однонитевых молекул такова, что их концы взаимно комплементарны
и поэтому способны взаимодействовать друг с другом, образуя структуру, называемую
"ручкой сковородки" (рис. 2, а). Регенерация утерянного по какой-либо причине
3'-конца цепи (рис. 2, б ) большой проблемы для вируса не представляет, поскольку
структура полноразмерной ДНК может быть восстановлена в результате копирования
5'-конца этой же молекулы (рис. 2, в).
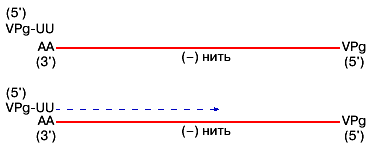
Возможна регенерация концевых нуклеотидов генома и у пикорнавирусов (рис. 3).
К 5'-концу вирусной РНК, на котором находятся два уридиловых остатка (UU), ковалентно
присоединен вирусный белок, называемый VPg. Синтез цепи вирусной
РНК на матрице комплементарной (-) нити начинается при участии нуклеотид-белкового
комплекса: к затравочному белку VPg сначала нематричным способом присоединяется
динуклеотид UU, который соответствует 5'-концу вирусной РНК. Удаление одного-двух
концевых остатков уридиловой кислоты из РНК некоторых пикорнавирусов не лишает
их инфекционности, поскольку эти нуклеотиды все равно приносят с собой VPg-UU.
Ясно, что у потомства укороченных РНК воссоздается нормальная концевая последовательность.
|
| Рис. 1. Полярность матричной (сплошная линия) и вновь синтезируемой (пунктир)
цепей нуклеиновых кислот |
Особый класс вирусной помехоустойчивости связан с дублированием генов или
(значительно чаще) с дублированием функций генов. Например, у многих аденовирусов
есть два участка, кодирующие так называемые вирус-ассоциированные РНК (VA1 и
VA2). Эти похожие (хотя и неидентичные), относительно низкомолекулярные РНК
не кодируют белков, но препятствуют подавлению белкового синтеза, которое могло
бы произойти в результате защитных мероприятий зараженной клетки. Поскольку
обе вирус-ассоциированные РНК действуют сходным образом, инактивация одного
из VA-генов не приводит к серьезным неприятностям. Случай дупликации (точнее,
трипликации) генов известен и у пикорнавирусов: у вируса ящура есть три весьма
сходных гена, кодирующие VPg. Вирус остается жизнеспособным (хотя и ослабленным)
при сохранении хотя бы одного из них.
|
Рис. 2. Схема, объясняющая возможность "регенерации" концов аденовирусной
ДНК. Схема регенерации 3'-концевого участка цепи ДНК. Комплементарные концы
промежуточной полноразмерной однонитевой ДНК взаимодействуют друг с другом
и образуют "ручку сковородки" (а). При делеции 3'-концевого участка (б )
он может быть полностью восстановлен в результате копирования 5'-конца той
же цепи (в) |
Чаще можно наблюдать частичное дублирование функций вирусных белков. Чем крупнее
геном, тем больше примеров такого рода. У аденовирусов весьма распространен
альтернативный сплайсинг, в результате которого один ген обеспечивает образование
нескольких молекул мРНК, различающихся по расположению и длине участков (интронов),
удаляемых из этих молекул в процессе созревания (посттранскрипционного
процессинга). Соответственно образуется несколько белков, которые в некоторых
случаях близки по структуре (например, различаются наличием дополнительных доменов).
Родственные белки могут проявлять схожую (хотя и неодинаковую) биологическую
активность. Поэтому мутации в некоторых местах данного гена, инактивирующие
один из белков, могут остаться без последствий для другого белка этого же семейства
(например, если они попадут в интрон соответствующей РНК). В результате функция,
контролируемая этим геном, будет нарушена лишь частично (если вообще нарушена)
и вирус не потеряет жизнеспособности.
В приведенном примере белки со сходной функцией были хотя и разными, но сходными.
Бывает и частичное перекрывание функций совсем неродственных вирусных белков.
Например, несколько разных аденовирусных белков препятствуют развитию апоптоза
(программируемой смерти), который индуцируют другие белки этого вируса. Помимо
всего прочего перекрывание функций может рассматриваться как страховка против
возможных генетических повреждений.
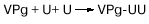
|
Рис. 3. Схема инициации полиовирусной РНК (пунктир) на комплементарной (-)
матрице при участии VPg-UU |
У аденовирусов, как и у других вирусов с крупным геномом, наряду с абсолютно
обязательными (эссенциальными) генами, без участия которых репродукция невозможна,
есть и такие, которые требуются либо для интенсификации какого-либо процесса,
либо только при особых условиях. В качестве примера приведем так называемую
область Е3, захватывающую около 10% генома и кодирующую несколько белков, помогающих
преодолевать защитные функции макроорганизма. Серьезные травмы этого участка
ДНК (например, делеции или замена чужеродными генами) для вируса
несмертельны.
Рассмотренные свойства, придающие вирусам устойчивость, относятся к числу тех,
которые выработаны в процессе эволюции и имеются у всех (или большинства) представителей
данного вида. Для того чтобы их сохранить в потомстве, необходима высокая точность
воспроизведения (репликации) генетического материала.
Далее...
Написать комментарий
|
