|
Клетка как архитектурное чудо.
I. Живые нити
Ю. М. ВАСИЛЬЕВ
Московский государственный университет им. М.В. Ломоносова
ВВЕДЕНИЕ
Каждый знает, что наш организм есть федерация огромного множества отдельных
клеток. Однако мы часто недооцениваем тот простой факт, что каждая из этих клеток
- сложный индивидуум, обладающий собственными принципами поведения. Если не
понять эти принципы, нельзя разобраться во взаимодействиях клеток в организме.
Изучать поведение отдельных клеток лучше всего, пользуясь методом клеточных
культур, то есть выделяя отдельные клетки из организма и помещая их в сосуд
с питательной средой. Если наблюдать эти клетки под микроскопом и фиксировать
их поведение на кино- или видеопленке, то легко убедиться в том, что каждая
клетка в такой культуре живет самостоятельной сложной жизнью: прикрепляется
ко дну сосуда и ползает по этому дну (подложке), меняя свою форму и направление
движения, выбрасывая и втягивая отростки. Внутри клеток отдельные пузырьки-органеллы
все время движутся. Долго казалось, что разобраться в механизмах этого сложного
поведения клеток и их частей почти невозможно.
Замечательное достижение последних десятилетий - открытие и исследование системы
структур, ответственных за подвижную архитектуру клетки, за ее движения и форму.
Этой системой в клетках эукариот оказался цитоскелет - система
белковых нитей, наполняющих цитоплазму (рис. 1). В этой статье я попытаюсь кратко
рассказать о том, как организован цитоскелет, каковы основные типы его конструкций.
ПОЛИМЕРИЗАЦИЯ И ДЕПОЛИМЕРИЗАЦИЯ НИТЕЙ - ОСНОВА ДИНАМИКИ ЦИТОСКЕЛЕТА
Цитоскелет состоит из трех основных типов нитей, образующих три системы: микрофиламенты,
микротрубочки и промежуточные филаменты. Каждый тип нитей состоит из одного-двух
основных белков: микрофиламенты - из актина, микротрубочки - из тубулина, промежуточные
филаменты - из специальных белков, различных в разных тканях: кератинов - в
эпителиях, десмина - в мышцах, виментина - в тканях внутренней среды (соединительной
ткани, хряще, кости и др.), белков нейрофиламентов - в нейронах.
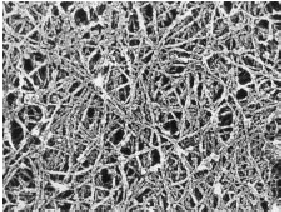
|
Рис. 1. Сеть актиновых микрофиламентов в цитоплазме культивируемой клетки
(фибробласта).
Микрофотография с трехмерной реплики цитоскелета под электронным микроскопом.
Увеличение 45 000i. Уменьшение при печати в 1.6 раза. Препарат Т.М. Свиткиной.
|
Разумеется, белки цитоскелета, как и любые белки клетки, закодированы в ДНК
и синтезируются на рибосомах. Клетка может менять набор синтезируемых
белков. Однако конструкция цитоскелета может быстро меняться даже без синтеза
новых молекул, за счет полимеризации и деполимеризации нитей. Отдельные молекулы,
мономеры, растворенные в цитоплазме клетки, способны соединяться, полимеризоваться
в нити соответствующего типа. Новые мономеры могут присоединяться к концам нити,
удлиняя ее. Полимеризация обратима: мономеры могут отделяться от концов нити,
которая при этом укорачивается и может исчезнуть совсем. В клетке все время
идет обмен между нитями и раствором мономеров в цитоплазме (рис. 2). Во многих
клетках примерно половина молекул актина и тубулина находится в виде мономеров
в цитоплазме и половина входит в состав актиновых нитей, микрофиламентов или
микротрубочек. Локальные условия полимеризации могут часто меняться. Поэтому
одна и та же нить может то укорачиваться, то удлиняться (см. рис. 2).
Клетка регулирует стабильность нитей цитоскелета, присоединяя к ним специальные
белки, которые меняют скорость полимеризации и деполимеризации мономеров. Поэтому
нить, состоящая из одного и того же мономера, может иметь очень разную продолжительность
жизни. Например, индивидуальные микротрубочки, входящие в состав жгутика или
реснички, обычно живут много часов и дней. Напротив, каждая микротрубочка митотического
веретена, состоящая из того же тубулина, живет в среднем лишь несколько минут.
Микротрубочки веретена все время растут и распадаются, одни микротрубочки заменяются
другими. Между тем само веретено, то есть совокупность микротрубочек, идущих
от полюсов к хромосомам и экватору клетки, сохраняется в течение всего митоза,
лишь постепенно меняя свою тонкую структуру. Уже в середине митоза веретено
состоит из иных микротрубочек, чем в его начале. Пример с веретеном иллюстрирует
общий принцип работы большинства цитоскелетных систем, названный принципом динамической
нестабильности: отдельные нити в системе могут появляться и исчезать в результате
полимеризации-деполимеризации, и поэтому детальное строение системы постоянно
меняется, но, несмотря на это, общий план организации системы может сохраняться.
Разберем теперь, как проявляется динамическая нестабильность в работе каждой
из трех цитоскелетных систем.
СИСТЕМА МИКРОФИЛАМЕНТОВ
Мономеры актина полимеризуются в микрофиламенты диаметром около 6 - 7 нанометров
(1 нм = 10- 9 м). Микрофиламенты полярны: их концы неодинаковы. Полимеризация
микрофиламента на одном конце, называемом плюс-концом, идет легче, чем на другом,
минус-конце. Полимеризация и деполимеризация молекул регулируются разными актинсвязывающими
белками. Некоторые из таких белков присоединяются к одному концу нити, блокируя
на этом конце полимеризацию и деполимеризацию, тогда рост и укорочение микрофиламента
идут лишь на другом конце, не закрытом блокирующим белком. Некоторые специальные
белки соединяют несколько мономеров в "зачаток" нити, вызывают нуклеацию нового
микрофиламента. В дальнейшем такие нити растут в одну сторону, обычно в сторону
плюс-конца. Специальные белки могут присоединяться к бокам нескольких микрофиламентов.
При этом одни белки связывают микрофиламенты в сети, другие - в пучки.
Особую роль среди актинсвязывающих белков играют миозины, так как они могут
двигаться по микрофиламенту. В настоящее время известна структура свыше 80 вариантов
молекул миозинов. У всех миозинов молекула состоит из трех частей: головки,
шейки и хвоста. Головка способна присоединяться к боку актинового микрофиламента,
и если снабжать эти головки поставляющим химическую энергию веществом - АТФ,
то головка движется вдоль микрофиламента, от плюс- к минус-концу, перескакивая
с одного мономера на другой. Этот процесс - основа очень многих движений в клетке.
Характер этих движений во многом зависит от структуры того миозина, который
его осуществляет, от того, каковы у этой молекулы головки и хвосты. Например,
молекула обычного миозина из поперечнополосатых мышц человека, так называемого
миозина II, имеет длинный хвост. Переплетаясь хвостами, эти молекулы образуют
миозиновые филаменты с торчащими наружу многими головками (рис. 3). В мышечной
клетке очень стабильные актиновые микрофиламенты расположены параллельно друг
другу на фиксированных расстояниях друг от друга и от миозиновых филаментов,
помещающихся между ними. Прикрепляясь к актиновым филаментам, головки миозиновых
нитей движутся вдоль этих филаментов, и это скольжение - основа всех мышечных
движений (см. рис. 3). У других миозинов, например у так называемых миозинов
I, хвосты очень короткие. Поэтому такие миозины, в отличие от миозинов II, переплетаться
хвостами и образовывать филаменты не могут. Вместо этого молекулы некоторых
миозинов I могут поодиночке прикрепляться своими короткими хвостами к мембранам
разных органелл (например, митохондрий, лизосом и др.). Если головка той же
молекулы одновременно прикрепится к актиновой нити, то она может двигать органеллу
вдоль этой нити (см. рис. 3).
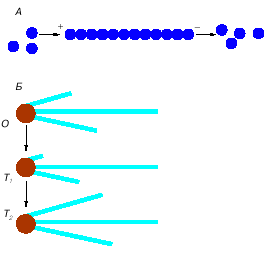
|
Рис. 2. Динамичность нитей цитоскелета.
А - микрофиламент, полимеризующийся из мономеров (синие кружки) на плюс-конце
и деполимеризующийся на мономеры на минус-конце.
Б - динамическая нестабильность системы из трех микротрубочек (параллельные
прямые), полимеризующихся из одного центра (кружок). О, Т1 , Т2 - последовательные
моменты времени. Верхняя микротрубочка сильно укоротилась в интервале О
- Т1 и вновь выросла в интервале Т1 - Т2 . Нижняя микротрубочка: укорочение
(О - Т1), затем рост (Т1 - Т2). Здесь и на рис.3 - 6 все структуры изображены
сугубо схематически, без соблюдения точности деталей. |
Комбинируя стандартные актиновые микрофиламенты с различными миозинами и другими
актинсвязывающими белками, клетка строит самые различные структуры, отличающиеся
по архитектуре и подвижности. Мы уже упоминали об одной из таких структур -
миофибрилле, образующейся в высоко специализированных мышечных клетках. Так
как в мышце все нити строго параллельны друг другу, то их скольжение и сокращение
всей мышцы идет в одном направлении и мышца может развить большое напряжение.
У большинства других клеток, например в клетках соединительной ткани (фибробластах),
клетках эпителия, лейкоцитах и других клетках, большая часть микрофиламентов
образует другую структуру - актиновый кортекс, располагающийся под мембраной.
Кортекс, подобно миофибрилле, может сокращаться за счет взаимодействия актиновых
микрофиламентов с миозиновыми молекулами. Однако, в отличие от миофибриллы,
в кортексе микрофиламенты далеко не всегда параллельны друг другу, часто они
образуют сложные сети. Поэтому сжатие кортекса идет обычно в нескольких направлениях.
Кроме того, в кортексе, в отличие от миофибриллы, микрофиламенты очень динамичны;
кортекс все время обновляется и перестраивается путем полимеризации-деполимеризации
нитей. Если средняя продолжительность жизни микрофиламента в миофибрилле более
7 дней, то в кортексе лейкоцита - всего лишь 5 с.
Основным и очень важным типом перестроек кортекса являются псевдоподиальные
реакции: выбрасывание, прикрепление и сокращение псевдоподий. Рассмотрим подробнее
эти реакции. При выбрасывании псевдоподии на поверхности клетки очень быстро,
в течение нескольких минут или даже секунд, образуется вырост цитоплазмы. Такой
вырост может иметь разную форму, например форму плоской пластинки (ламеллоподия),
узкого цилиндра (филоподия) или просто шаровидного пузыря. Внутреннее строение
всех типов псевдоподий просто: они часто не содержат никаких структур, кроме
кортикальных микрофиламентов. При этом в ламеллоподиях эти микрофиламенты образуют
густую уплощенную сеть, а в пузырях - менее упорядоченный слой под мембраной
(рис. 4). Если микрошприцем инъецировать в клетку раствор мономеров актина,
помеченных флуоресцирующей краской, а затем наблюдать такую клетку в флуоресцентном
микроскопе, где краска ярко светится, то можно видеть, что микрофиламенты из
меченых мономеров появляются раньше всего именно в псевдоподиях. Таким образом,
псевдоподии являются местом, где из мономеров полимеризуются микрофиламенты.
Вероятно, под мембраной в этих местах концентрируются какие-то белки, вызывающие
полимеризацию новых микрофиламентов, но пока природу этих белков мы еще точно
не знаем.
Форма выпячивания может определяться тем, с какими белками свяжутся вновь возникшие
микрофиламенты. Это подтверждается недавними опытами Штосселя (Stossel T. Science,
1993. V. 260. P. 1086). Он обнаружил, что клетки одной из линий клеток в культуре
выпячивают на поверхности лишь шаровидные пузыри, но не ламеллоподии. Оказалось,
что в геноме этих клеток отсутствовал ген, кодирующий белок, который связывает
актиновые микрофиламенты в сеть. Специальными методами генной инженерии исследователи
ввели в клетки недостающий ген, и тогда клетки стали делать не пузыри, а уплощенные
ламелоподии. Таким образом, появление в актиновом кортексе одного дополнительного
белка направленно изменило архитектуру псевдоподий.
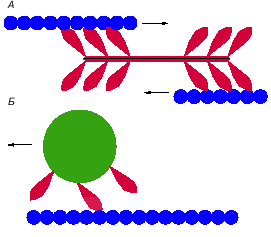
|
Рис. 3. Взаимодействия актиновых микрофиламентов (нити из синих кружков)
с миозинами (красные структуры).
А - схема движений в миофибрилле мышцы. Молекулы миозина II соединены длинными
хвостами в нить, из которой наружу торчат в разные стороны головки. Головки
миозиновых молекул движутся по двум параллельным актиновым нитям, вызывая
скольжение этих нитей в двух противоположных направлениях.
Б - схема движения органеллы (зеленый круг) вдоль микрофиламента при помощи
миозина I. Молекулы миозина I короткими хвостами прикреплены к мембране
органеллы, а концами головок - к актиновой нити. |
Поверхность конца выброшенной псевдоподии может прикрепиться к подложке, по
которой ползет клетка. При этом образуется место прочного контакта, где определенные
белки мембраны наружным концом молекулы соединяются с белками, прикрепленными
к подложке; внутренним концом та же молекула соединяется, через ряд промежуточных
звеньев, с актиновыми микрофиламентами псевдоподии.
Сразу после выбрасывания псевдоподия содержит актин, а миозин II проникает в
псевдоподию (диффундирует) из внутренней части клетки лишь несколько минут спустя.
Взаимодействие миозинов с актиновыми нитями вызывает сокращение псевдоподии.
Это сокращение может иметь разные последствия для клетки. Если псевдоподия не
прикреплена к подложке, она втягивается и исчезает. Напротив, если псевдоподия,
выброшенная на одном из краев клетки, успела прочно прикрепиться к подложке,
то сокращение ее смещает вперед все тело клетки. Повторяя псевдоподиальные реакции,
клетка ползет по подложке. Если много псевдоподий на разных краях клетки выбрасываются
и прикрепляются к подложке одновременно, то они, стремясь сократиться, натягиваются
и растягивают клетку в разные стороны, уплощая ее форму. Этот процесс называют
распластыванием.
Термин "псевдоподия" означает в переводе - ложная ножка. Это действительно ножка,
которая двигает клетку вперед по подложке. Вместе с тем это ножка особая: в
отличие от ноги человека псевдоподия может вырасти заново из тела клетки, образовать
свои мышцы, сократиться и исчезнуть за считанные минуты. Как мы видели, эволюция
псевдоподии является результатом серии сложных молекулярных реакций: полимеризации
актиновых нитей, присоединения к этим нитям других белков, связывающих их в
сети и вызывающих их перемещение, а также связывания нитей с белками мембраны.
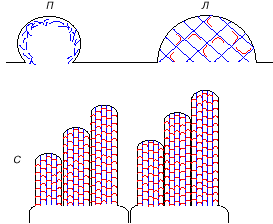
|
Рис. 4. Выросты поверхности клетки, образуемые актиновыми микрофиламентами
(синие линии).
А - два варианта псевдоподий. П - пузырь, где под мембраной клетки имеется
слой коротких микрофиламентов, не имеющий упорядоченной организации. Л -
ламеллоподия - пластинчатый вырост, где микрофиламенты соединены в упорядоченную
сеть молекулами специального актин-связывающего белка (изогнутые утолщенные
линии).
Б - стереоцилии на поверхности двух соседних волосковых клеток улитки внутреннего
уха. Вертикальные параллельные актиновые микрофиламенты в каждой стереоцилии
связаны друг с другом и с клеточной мембраной молекулами миозинов и других
актинсвязывающих белков (горизонтальные красные линии).
В одной клетке разные ряды стереоцилий имеют разную строго фиксированную
длину. В соседних клетках стереоцилии соответствующих рядов (среднего и
правого) также имеют разную фиксированную длину. |
Псевдоподии - лишь один из многих типов отростков, содержащих актиновые микрофиламенты.
Расскажем еще об одном виде весьма важных специализированных отростков, так
называемых стереоцилиях (см. рис. 4). Эти отростки располагаются пучками на
верхней поверхности волосковых клеток улитки внутреннего уха, клеток, отвечающих
за восприятие звуков. Стереоцилии наполнены от основания до верхушки параллельными
друг другу стабильными актиновыми микрофиламентами; между ними и мембраной имеются
молекулы других белков, в том числе специальных миозинов. В каждой клетке все
стереоцилии и их микрофиламенты имеют строго определенные размеры; "допуск"
вариаций такой длины не более 5%. На поверхности каждой клетки они располагаются
подобно трубам органа, рядами "по росту": самые короткие в переднем ряду, а
самые длинные - в заднем (см. рис. 4). При этом длина и ширина стереоцилии самого
длинного ряда в разных волосковых клетках закономерно меняются от клетки к клетке
вдоль поверхности улитки (см. рис. 4). Такая точность размеров стереоцилий очень
важна, так как разные клетки улитки активируются звуками разной частоты и амплитуды:
низкие звуки воспринимаются клетками на одном конце улитки, высокие - на другом.
Полагают, что звуковая волна вызывает колебания стереоцилий, притом на различные
частоты звука резонируют стереоцилии разной длины и, следовательно, активируются
разные клетки.
Мы пока еще не знаем, как в эмбриональном периоде создается замечательная "музыкальная"
архитектура стереоцилий; это увлекательная проблема для исследователей. Совсем
недавно было сделано еще одно неожиданное открытие. Существует особая наследственная
глухота и слепота, так называемый синдром Ашера; одно из его проявлений - дегенерация
волосковых клеток уха. Большая группа исследователей (Weil et al. Nature, 1995.
V. 374. P. 60.) показала, что основой болезни является мутация, инактивирующая
один из особых миозинов, так называемый миозин VIIА. Что именно делает этот
миозин в волосковых клетках, как он участвует в реакциях стереоцилии на звук,
мы пока не знаем. Еще менее понятно, почему тот же миозин необходим для восприятия
света клетками сетчатки глаза, почему без одного этого белка цитоскелета человек
становится не только глухим, но и слепым. Мы еще и еще раз убеждаемся, как велико
разнообразие конструкций, которые клетка умеет строить на основе одного основного
элемента цитоскелета - актиновых микрофиламентов.
Далее...
Написать комментарий
|
