Бордунов Сергей Иванович
Автореферат диссертации на соискание ученой степени кандидата геолого-минералогических наук
|
содержание |
4.1. Описание разрезов
о. Карагинский
Разрез неогена о. Карагинского является опорным для Восточной Камчатки (Гладенков, 1972; Гладенков и др., 1992). В опорном разрезе снизу вверх залегают следующие отложения:
Миоцен. Нижний - средний миоцен. Пестроцветная свита.
Пестроцветная свита с небольшим размывом залегает на отложениях свиты песчаников с Laternula и сложена в основном тефрогенными песчаниками, гравелитами, алевролитами, глинистыми и кремнистыми (диатомовыми) алевролитами и аргиллитами общей мощностью 380-425 м. В составе свиты выделяется четыре толщи (толщи 10-13).
Нижняя толща 10 (мощностью 110-115 м) представлена зеленовато-серыми песчаниками (60-75 м), содержащими обильные карбонатные конкреции. Они подстилаются пачками темно-серых до черных гравелитов и песчаников (39 м).
Вышележащая толща 11 (50 м) отделена сбросом. Она представлена темными зеленовато-серыми алевролитовыми и диатомовыми аргиллитами с частыми тонкими прослоями песчаников.
Толща 12 (130 м) имеет флишоидное строение и сложена частым и грубым переслаиванием алевролитов, диатомовых алевролитов, аргиллитов и слоев песчаников, гравийных песчаников, песчаников с гальками и валунами.
Толща 13 (100-130 м) представлена глинистыми и диатомовыми алевролитами со слоями туфогенных гравийно-галечных песчаников в нижней части.
Средний - верхний миоцен. Свита мыса Плоского
В свите мыса Плоского выделено шесть толщ, из которых первые четыре объединяются в нижнюю, а две остальные - в верхнюю подсвиты. Нижняя толща 1 сложена зеленовато-серыми песчаниками (более 20-25 м). Толща 2 представлена туфоалевролитами и диатомовыми туфоалевролитами (150-190 м). Толща 3 включает переслаивающиеся туфы, глинистые туфодиатомиты (40-60 м). Толща 4 (до 150 м) сложена сравнительно однородными туфоалевритовыми и глинистыми туфодиатомитами с прослоями тефроидов, туфов и с карбонатными конкрециями. Толща 5 (130-140 м) резко отличается от подстилающих отложений. Она представлена чередованием пачек алеврито-глинистых диатомитов и ритмично переслаивающихся тефрогенных пород: песчаников, алевролитов и аргиллитов. Залегающая выше толща 6 (190-210 м) более однородна и сходна с толщей 4.
Верхний миоцен. Юнюньваямская свита
Юнюньваямская свита согласно, но с размывом залегает на свите мыса Плоского. В ее составе различаются три толщи (7-9). Нижняя толща 7 сложена гравелитами и песчаниками (20 м). Толща 8 состоит из диатомовых туфоалевролитов и туфодиатомитов (170-185 м). Толща 9 (123 м) представлена туфопесчаниками и конгломератами с пачкой базальтовых тефроидов и туфобрекчий в основании.
Плиоцен. Лимимтэваямская свита.
Лимимтэваямская свита залегает на подстилающей ее юнюньваямской свите с размывом, но, видимо, согласно. В составе лимимтэваямской свиты выделено три толщи. Нижняя толща 10 соответствует нижней подсвите. Она представлена песчаниками и конгломератами (95,5-109 м). Верхняя подсвита начинается толщей 11 (60-62 м), которая включает алевролитовые туфодиатомиты и туфоалевролиты. Сменяющая ее толща 12 (103 м) литологически схожа. В лимимтэваямской свите выделяется 14 мощных маркирующих прослоев вулканических пеплов.
Усть-лимимтэваямская свита. Залегает на подстилающих толщах с некоторым размывом, но с видимым согласием, и представлена толщей 13 (77-80 м). Толща 13 (туфопесчаники и туфоалевролиты) включает пачки от 1 до 7.
Тусатуваямские слои. Толща 14 (12,5 м) - конгломераты, с размывом, залегающие на туфоалевролитах толщи 13.
п-в Камчатский
Наиболее полный разрез ольховской свиты вскрывается по долине р. Мутной, где ее выходы прослеживаются от устья на 4 км вверх по течению.
Плиоцен - эоплейстоцен. Ольховская свита.
Нижняя подсвита. Толща 1 сложена конгломератами с прослоями песчаников и глин. Толща 2 представлена песчаниками и алевролитами. Верхняя подсвита. Толща 3 состоит из конгломератов гравийных и галечных,
перекрывается с размывом песками и галечниками верхнего плейстоцена и голоцена.
В разрезах ольховской свиты присутствует граница плиоцена и плейстоцена (Петров, 1982). Таким образом, они дополняют Карагинский разрез, где граница неогена и квартера достоверно не установлена. В совокупности изученные разрезы составляют практически полную последовательность неогеновых отложений. Это позволяет составить представление о стратиграфической последовательности комплексов бентосных фораминифер на протяжении всего неогена.
4.2. Биостратиграфическое расчленение неогена по фораминиферам (слои с фауной)
С целью детального расчленения неогеновых отложений Карагинского разреза были изучены фораминиферы из всех перечисленных выше свит. Анализ комплексов фораминифер позволил выявить их довольно четкую последовательную смену. На этом основании впервые выделены биостратиграфические подразделения в ранге слоев с фауной.
Нижняя граница каждого такого подразделения обычно устанавливалась по появлению нескольких новых видов, чему соответствовала определенная перестройка качественной и количественной структуры комплексов фораминифер (рис. 1). Обычно границы слоев с фораминиферами смыкаются в разрезе. Если отложения между соседними слоями не содержали раковин фораминифер, то они выделялись как интерслои. За виды индексы в названии слоев обычно принимались два - один, характерный для этого слоя, и второй, доминирующий в нем, как правило, с достаточно определенным стратиграфическим положением и широким географическим распространением.
 | | Рис. 1. Стратиграфическое распространение характерных видов фораминифер в разрезе неогена о. Карагинского. Условные обозначения: цифры в кругах - номер слоев с фораминиферами, остальные обозначения см. рис. 2. |
Отложениях миоцена и плиоцена о. Карагинского расчленены на 21 подразделение в ранге слоев с фораминиферами. Все слои с фауной привязаны к литологическим пачкам или их частям (Гладенков и др., 1992). В плиоцен-эоплейстоцене п-ва Камчатского (ольховская свита) выделены 4 подразделения в ранге слоев с фораминиферами.
В стратиграфической последовательности это следующие слои:
О. Карагинский
Миоцен. Нижний миоцен. Пестроцветная свита.
1. Слои с Pseudoelphidiella subcarinata и Melonis tumiensis (литологическая пачка 1 по Гладенкову и др., 1992). Комплекс фораминифер включает 20 видов, преобладают Pseudoelphidiella subcarinata (Voloshinova) и Melonis tumiensis V.Kuznetzova. Встречены Discoislandiella curvicamerata (Voloshinova).
Нижний-средний миоцен.
2. Слои с Spirosigmoilinella compressa и Martinottiella communis (пачки 2-3). Преобладают Martinottiella communis d'Orbigny. Хорошими стратиграфическими реперами являются Haplophragmoides kakertensis Serova, Spirosigmoilinella compressa Matsunaga.
3. Слои с Haplophragmoides tortuosus и Asanospira carinata (пачка 4). Имеют довольно обедненный комплекс фораминифер. Доминируют Asanospira carinata (Cushman et Renz), Globobulimina pacifica Cushman и Haplophragmoides kakertensis Serova.
Средний миоцен. Свита мыса Плоского
4. Слои с Elphidiella semiinvoluta и Discoislandiella umbonata (пачки 1 и 2) характеризуются более мелководным комплексом фораминифер. Количественно преобладают Cassandra excavata (Voloshinova) и Discoislandiella umbonata (Voloshinova). В акцессорной группе видов отмечена Elphidiella semiinvoluta Voloshinova.
5. Слои с Cribrostomoides sakhalinensis и Haplophrаgmoides indentatus (нижняя часть пачки 3). Характеризуются резкой сменой состава, так как комплекс образован почти исключительно агглютинирующими видами. Преобладают Haplophragmoides impressus Voloshinova, Cyclammina postpilvoensis V.Kuznetzova, им сопутствуют менее многочисленные Cyclammina praecancellata Voloshinova, Cribrostomoides sakhalinensis Voloshinova, Asanospira carinata (Cushman et Renz). Появляются такие реперные виды для среднемиоценовых отложений, как Plectina nipponica Asano, Haplophragmoides indentatus Voloshinova.
6. Слои с Dorothia paupercula и Gaudryina quadrangularis (верхняя часть пачки 3 и пачка 4). Отличается массовым развитием Gaudryina quadrangularis Bagg. Доминируют Gaudryina quadrangularis, Asanospira carinata. Встречается значительное количество Haplophragmoides indentatus.
Средний-Верхний миоцен.
7. Слои с Cribroelphidium subglobosum и Cassilamellina californica (пачки 5 и 6). Для комплекса фораминифер этих слоев характерно преобладание секреционных форм над агглютинирующими. Доминирует Cassilamellina californica (Cushman et Hughes), ему сопутствуют Cribroelphidium subglobosum (Voloshinova), Globobulimina pacifica Cushman, Discoislandiella smechovii (Voloshinova).
8. Слои с Budashevaella laevigata и Haplophragmoides indentatus (пачки 7-12). Агглютинирующие формы резко преобладают над секреционными. В массовом количестве встречаются Haplophragmoides indentatus, H. spadix V.Kuznetzova, Martinottiella communis.
Верхний миоцен.
9. Слои с Globobulimina pacifica и Cyclammina postpilvoensis (пачки 14-20) содержат обедненный комплекс фораминифер, состоящий всего лишь из четырех видов: Haplophragmoides spadix, Martinottiella communis и виды индексы.
10. Слои с Elphidiella nabilensis и Cassandra excavata (пачка 21). Комплекс представлен фораминиферами с секреционной стенкой. Количественно преобладают Elphidiella sp., Cassandra excavata (Voloshinova) и Elphidiella nabilensis Voloshinova et Borovleva.
11. Слои с Globobulimina pacifica и Cyclammina praecancellata (пачка 27). Комплекс обедненный, включает только три вида: виды индексы и Haplophragmoides impressus. Количественно преобладают Cyclammina praecancellata, их крупные раковины можно наблюдать в образцах породы невооруженным глазом.
Юнюньваямская свита
12. Слои с Globobulimina pacifica и Haplophragmoides impressus (пачки 1-5) характеризуются обедненным комплексом фораминифер, в котором доминируют агглютинирующие формы. Встречены Haplophragmoides impressus, Cyclammina praecancellata, Martinottiella communis, Globobulimina pacifica.
13. Слои с Quinqueloculina sakhalinica и Cribroephidium vulgare (пачки 6-9) содержит набор мелководных видов фораминифер. Количественно преобладают Cribroelphidium vulgare (Voloshinova), Haplophragmoides impressus. Выше в разрезе неогена о. Карагинского непереотложенные агглютинирующие фораминиферы не встречаются.
Плиоцен. Лимимтэваямская свита
14. Слои с Elphidiella nagaoi и Е. oregonensis (пачки 5-10); Характерно массовое развитие Е. oregonensis (Cushman et Grant) и доминирование представителей рода Elphidiella.
15. Слои с Elphidiella arctica и Cibicides rotundatus (пачки 11-13). Преобладают Elphidiella hannai (Cushman et Grant) и Cibicides rotundatus Schedrina. Впервые в разрезе отмечены бореально-арктические виды Elphidiella arctica (Parker et Jones) и Cribroelphidium subarcticum (Cushman).
16. Слои с Cribroelphidium vulgare и Islandiella laticamerata (пачка 14). В массовом количестве встречаются Retroelphidium clavatum (Cushman),
Islandiella laticamerata (Voloshinova). Основу комплекса составляют бореальные и бореально-арктические виды.
17. Слои с Lagena gracilis и Nonionella globosa (пачка 15). Преобладает Islandiella laticamerata. Менее многочисленны раковины родов Buccella, семейства Elphidiidae. Встречаются единичные планктонные фораминиферы Globigerina woodi Jenkins.
18. Слои с Islandiella japonica и Bolivina pseudopunctata (пачки 16-19 толщи 11 и пачки 20-21 толщи 12). Для комплекса фораминифер характерно большое видовое разнообразие (34 вида). Преобладают Islandiella japonica (Asano et Nakamura) и Retroelphidium clavatum. Отмечены также единичные представители планктонных фораминифер Globigerina falconensis Blow и Globigerina sp. Впервые появляются Epistominella pacifica (Cushman), Cribroelphidium bartletti (Cushman), Buccella inusitata Andersen, Trifarina kokozuraensis (Asano).
19. Слои с Retroelphidium hughesi и Epistominella pacifica (пачки 22-27). Преобладают представители рода Buccella: В. citronea Leonenko, B. pseudofrigida Leonenko, В. hannai arctica Voloshinova. Также многочисленны Retroelphidium clavatum, R. hughesi (Cushman et Grant), Epistominella pacifica, Cribroelphidium bartletti. В некоторых образцах встречены единичные батиальные Melonis pompilioides (Fichtel et Moll). Встречен один вид планктонных фораминифер Globigerina uvula Ehrenberg.
Усть-лимимтэваямская свита.
20. Слои с Cassandra inflata и Buccella pseudofrigida (пачки 28-34). Количественно преобладают Buccella pseudofrigida. Характерно появление новых видов: Buccella conica Voloshinova, Cassandra inflata (Gudina), Planocassidulina kaziwazakiensis (Husezima et Maruhasi), которые встречаются и в четвертичных отложениях северо-бореальных областей (Гладенков и др., 1994, Гудина и др., 1968).
Тусатуваямские слои.
21. Слои с Hyanesina orbicularis и Globulina glacialis. Преобладают Retroelphidium clavalum, Buccella citronea. Характерно появление группы видов, широко распространенных в четвертичных отложениях бореальных и бореально-арктических районов: Hyanesina orbicularis (Brady), Globulina glacialis Cushman et Ozawa.
П-в Камчатский. Плиоцен-эоплейстоцен. Ольховская свита
22. Слои с Cribroelphidium subarcticum и Retroelphidium hughesi (литологические пачки 1-15 по Петрову, 1982). Количественно преобладают Retroelphidium hughesi и Cribroelphidium bartletti. Важную роль в комплексе играют аркто-бореальные виды Hyanesina orbicularis (Brady), Globulina
glacialis Cushman et Ozawa, Cribroelphidium subarcticum (Cushman), Buccella hannai arctica, Retroelphidium clavatum.
23. Слои с Siphonoperta agglutinata и Discoislandiella subarctica (нижняя часть пачки 16). Характерно массовое развитие Discoislandiella subarctica sp.nov. В значительном количестве встречаются Retroelphidium hughesi, исландиеллиды Cassilamellina californica (Cushman et Hughes), Islandiella islandica (Norvang).
24. Слои с Uvigerina peregrina и Epistominella pacifica (верхняя часть пачки 16). Характерны Cribroelphidium batiale Troitskaja, Buliminella elegantissima (d'Orbigny), Sigmomorphina lautenshlagerae Kusina, Neogloboquadrina pachyderma (Ehrenberg), Uvigerina peregrina Cushman. Количественно преобладают Discoislandiella subarctica и Epistominella pacifica (Cushman). Данный комплекс имеет наиболее глубоководный облик в изучаемом разрезе.
25. Слои с Retroelрhidium hughesi и Cribroelphidium bartletti (пачка 18). Содержит достаточно мелководный комплекс фораминифер. Доминируют Retroelphidium hughesi и Cribroelphidium bartletti. В значительном количестве присутствуют Epistominella pacifica, Nonionella labradorica, Retroelohidium clavatum.
Выделенные слои с фораминиферами по составу бентосных комплексов были сопоставлены с существующими данными по комплексам фораминифер стратиграфических подразделений неогена соседних районов Северо-Западной Пацифики. Наряду с этим, использование данных физических методов позволило более обоснованно подойти к датированию изучаемых толщ (Гладенков и др., 1992). Так данные по трековому датированию вулканического стекла из пепловых прослоев определяют возраст пачки 20 толщи 5 свиты мыса Плоского в 12,13±0,83 млн лет (рис. 2); пачки 23 толщи 6 свиты мыса Плоского 11,05±1,12 млн лет; пачки 3 толщи 8 юнюньваямской свиты 6,95±0,58 млн лет; пачки 5 толщи 8 юнюньваямской свиты 6,22± 0,57 млн лет. В это же время K-Ar датировка обломков базальтов из туфобрекчий пачки 8 толщи 9 юнюньваямской свиты дает возраст для этой толщи равный 9,4±0,4 млн лет.
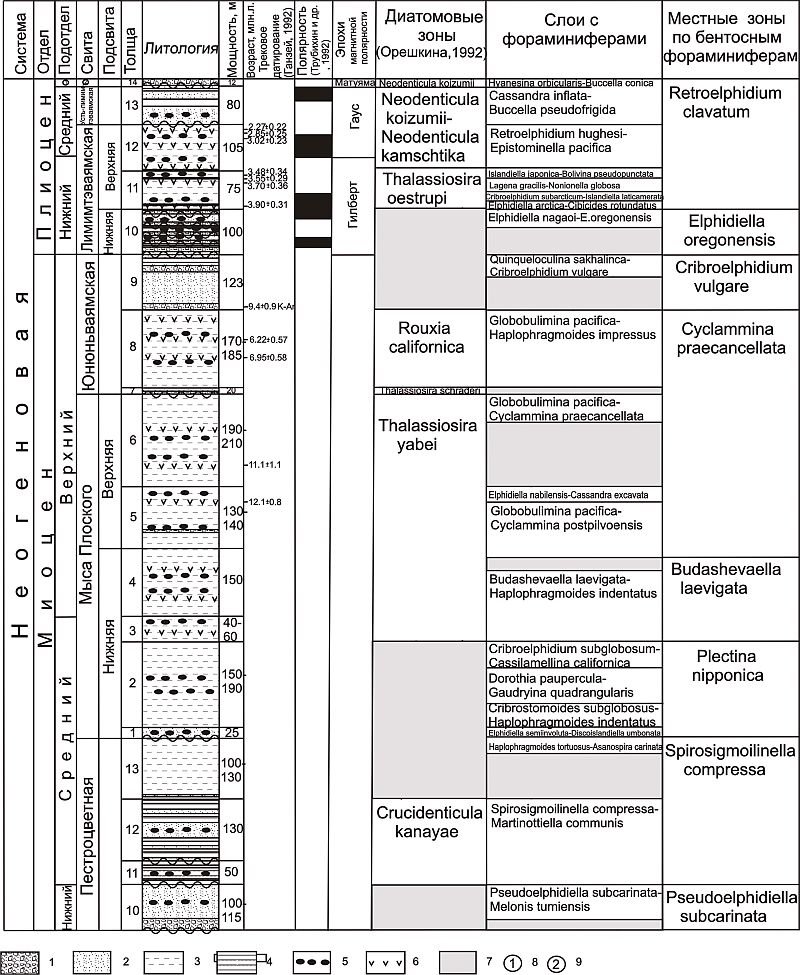 | Рис. 2. Стратиграфическое расчленение разреза неогена о. Карагинского по фораминиферам.
Условные обозначения: 1 - конгломераты и гравелиты; 2 - песчаники и туфопесчаники; 3 - алевролиты и туфоалевролиты; 4 - аргиллиты и туфоаргиллиты; 5 - карбонатные конкреции; 6 - прослои туфов; 7 - отсутствие фауны; 8 - верхний; 9 - тусатуваямские слои |
Палеомагнитные характеристики толщи 9 юнюньваямской свиты соответствуют верхней части эпохи Гильберт с возрастным интервалом 5,41-4,79 млн лет. По диатомовым водорослям отложения толщ 11-12 пестроцветной свиты коррелируются с зоной Crucidenticula kanayae (17,9-16,0 млн лет) зональной шкалы Северо-Тихоокеанской области (Орешкина, 1993); осадки верхней части свиты мыса Плоского (толщи 3-6) уверенно сопоставляется с зоной Thalassiosira yabei (12,9-11,6 млн лет); толщи 7-8 юнюньваямской свиты - с зоной Rouxia californica (7,3-6,6 млн лет); толща 9 этой свиты - с зоной Neodenticula kamtschatica (6,6-5,1 млн лет). Данные по моллюскам из пестроцветной, мыса Плоского и юнюньваямской свит также указывают на их формирование, соответственно, с раннего по поздний миоцен. По палеомагнитным данным лимимтэваямская свита соотносится со знакопеременной частью эпохи Гильберт с возрастом 4,5-3,8 млн лет; усть-лимимтэваямская свита - с верхней частью эпохи Гильберт и нижней частью эпохи Гаусс в интервале 3,6-3,25 млн лет; тусатуваямские слои - с эпохой Матуяма в интервале 2,48-1,87 млн лет) и трековому датированию (3,90±0,31 млн лет для пачки 14 толщи 11 лимимтэваямской свиты; 3,70±0,31; 3,55±0,29; 3,48±0,34 млн лет для пачек 16-18 толщи 11 лимимтэваямской свиты; 3,02±0,23 млн лет для пачки 23 толщи 12; 2,85 ± 0,25 млн лет для пачки 24 толщи 12; 2,27±0,22 млн лет для пачки 25 толщи 12). По диатомовым водорослям отложения толщи 10 лимитэваямской свиты сопоставляются с верхней частью зоны Neodenticula kamtschatica и нижней частью зоны Thalassiosira oestrupii; толща 11 - с зоной Thalassiosira oestrupii (5,1-4,3 млн лет); толщи 12 и 13 соответствуют зоне Neodenticula kamtschatica - Neodenticula koizumi (4,3-2,5 млн лет); тусатуваямские слои - зоне Neodenticula koizumi (2,5-1,89 млн л.). Данные по моллюскам из лимимтэваямской и усть-лимимтэваямской свит, а также тусатуваямских слоев указывают на формирование этих отложений с раннего по поздний плиоцен.
Возраст отложений ольховской свиты (слои 1-4) по палеомагнитным данным может быть определен как верхнеплиоценовый для слоев 1-2, и соответствует диатомовой зоне Neodenticula koizumi (2,5-1,89 млн лет), и как эоплейстоценовый для слоя 3 (диатомовая зона Actinocyclus oculatus) и нижнеплейстоценовый для слоя 4 (диатомовая зона Proboscia curvirostris) (Гладенков, 2007).
Полученные результаты по фораминиферам показали, в целом, хорошую сходимость с данными по другим группам фауны и флоры, а также с результатами физических методов.
4.3. Местные зоны и этапность развития неогеновых фораминифер Восточной Камчатки.
В результате анализа слоев с фауной и с учетом данных по составу и стратиграфическому распространению фораминифер в разрезах можно
наметить этапность в развитии неогеновой микрофауны в морских бассейнах Восточной Камчатки, которая отражается в определенной общности систематического состава и количественных соотношений таксонов в группах слоев (пачках) с фауной. Смена комплексов бентосных фораминифер соответствует, с одной стороны, эволюционному развитию фораминифер, а с другой - изменения палеообстановок (глубины бассейна, температуры вод, колебания уровня моря). Отмечаемые перестройки состава и структуры комплексов, тесно связаны с историей развития пришельфового палеобассейна Восточной Камчатки. Такие группы слоев были выделены по палеонтологическому и стратиграфическому критериям в виде биостратиграфических комплексных зон, а по масштабу их распространения - как местные зоны. В неогене Карагинского разреза выделяется 8 таких зон, которые прослеживаются и в смежных районах (табл. 1).
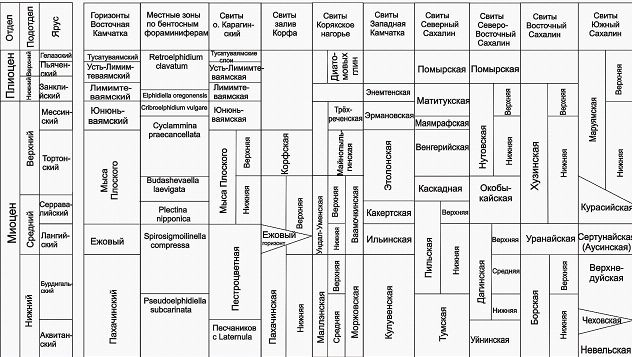 | | Таблица 1. Схема корреляции неогеновых отложений Восточной Камчатки со смежными регионами по бентосным фораминиферам. |
I. Зона Pseudoelphidiella subcarinata (включает слои с фауной 1). Выделяется в пределах толщи 10 (Гладенков и др., 1992) пестроцветной свиты. Нижняя граница устанавливается по появлению вида индекса. Характеризуется комплексом фораминифер со смешанным набором видов, в котором представлены как относительно мелководные секреционные фораминиферы Pseudoelphidiella subcarinata, Cribroelphidium crassum, Porosorotalia tumiensis, Discoislandiella curvicamerata, так и более глубоководные секреционные Melonis tumiensis, Melonis pompilioides, агглютинирующие Haplophragmoides laminatus, H. oblongus. Некоторые виды перешли из олигоцена. Состав комплекса отражает присклоновые условия с накоплением осадков турбидитного генезиса. Глубины бассейна соответствовали батиали.
Соответствует одноименной провинциальной зоне Юго-Западной Камчатки (Дмитриева, Фрегатова, 2004; Дмитриева, 2005). Зональный комплекс сходен с комплексами кулувенского горизонта Западной Камчатки, а также с моржовской и верхней подсвитой маллэнской свиты Хатырского прогиба, собольковской и нижней части гагаринской свит Анадырского прогиба, одянской свиты Магаданской скважины. Он обнаруживают сходство с комплексами уйнинского и дагинского горизонтов Северного Сахалина, невельского и углегорского горизонтов Центрального и Южного Сахалина. Зона соответствует нижней части пахачинского горизонта Восточной Камчатки. Нижний миоцен.
II. Зона Spirosigmoillina compressa (слои 2-3). Выделяется в пределах толщ 11-13 пестроцветной свиты. Нижняя граница устанавливается по появлению вида индекса и сопутствующего набора новых видов агглютинирующих фораминифер. Доминируют глубоководные агглютинирующие формы Spirosigmoilinella compressa, Haplophragmoides indentatus, Cyclammina pilvoensis, Martinottiella communis, Asanospira carinata и другие. На нижней границе зоны происходит резкая смена состава комплекса фораминифер. Состав комплекса фораминифер свидетельствует о резком углублении бассейна, вероятно, связанным как с начавшейся трансгрессией моря, так и его тектоническим прогибанием. Глубины соответствовали средней - нижней батиали.
Соответствует в полном объеме или частично зоне Martinottiella communis-Spirosigmoilinella compressa Юго-Западной Камчатки (Дмитриева, Фрегатова, 2004; Дмитриева, 2005), Alveolophragmium indentatus-Martinottiella communis о. Хоккайдо (Hanagata, Hiramatsu, 2005), зоне Spirosigmoilinella compressa северо-запада о. Хонсю (Inoue et al, 2008). Комплекс зоны Spirosigmoilinella compressa сопоставляются с комплексами автаткульской свиты Анадырского прогиба, верхов одянской свиты Магаданской скважины, нижней подсвитой ундал-уменской свиты Хатырского прогиба. По составу им близки и среднемиоценовые комплексы верхней части дагинского горизонта Северного Сахалина, уранайской свиты Пограничного прогиба и сертунайской (аусинской) свиты Центрального и Южного Сахалина. Зона соответствует верхней части пахачинского и ежовому горизонту Восточной Камчатки. На данный интервал приходится начало климатического оптимума конца раннего - начала среднего миоцена. Нижний - средний миоцен.
III. Зона Plectina nipponica (слои 4-7). Зона выделяется в пределах толщ 1-2 свиты мыса Плоского. Нижняя граница устанавливается по появлению вида индекса и ряда глубоководных агглютинирующих видов над базальной грубообломочной толщей 1. В комплексе преобладают агглютинирующие фораминиферы Haplophragmoides indentatus, Asanospira carinata, Cribrostomoides sakhalinensis. Часть видов переходит в данную зону из подстилающих отложений. Появляются новые виды агглютинирующих фораминифер Budashevaella laevigata, Cyclammina postpilvoensis, C. praecancellata. Присутствие относительно глубоководных видов Dorothia paupercula, Guadryina quadrangularis, Plectina nipponica свидетельствуют о значительной глубине палеобассейна (нижняя батиаль), существовавшего в районе исследований.
Соответствует части зоны Martinottiella communis-Spirosigmoilinella compressa Юго-Западной Камчатки (Дмитриева, Фрегатова, 2004; Дмитриева, 2005). Сопоставляется с какертским горизонтом Западной Камчатки, окобыкайским и курасийским горизонтами Сахалина. Зона соответствует нижней части горизонта мыса Плоского Восточной Камчатки. Средний миоцен.
IV. Зона Budashevaella laevigata (слои 8). Зона выделяется в пределах толщ 3-4 свиты мыса Плоского. Нижняя граница устанавливается по появлению вида индекса. Преобладают агглютинирующие формы с Haplophragmoides indentatus, H. kakertensis, Cribrostomoides sakhalinensis с постоянным присутствием Globobulimina pacifica. Характерными видами были агглютинирующие Budashevaella laevigata, Cyclammina postpilvoensis, Haplophragmoides impressus. Глубина палеобассейна составляла около 200-500 м. Состав комплекса фораминифер свидетельствует о холодноводных, обедненных кислородом условиях.
Возможно, соответствует части зоны Islandiella excavata-Haplophragmoides impressus Юго-Западной Камчатки (Дмитриева, Фрегатова, 2004; Дмитриева, 2005). Сопоставляется с верхней частью окобыкайского горизонта Северного Сахалина и нижней частью этолонского горизонта Западной Камчатки, а также низами майнопыльгинской свиты Корякского нагорья. Зона соответствует средней части горизонта мыса Плоского. Средний миоцен- - верхний миоцен.
V. Зона Cyclammina praecancellata (слои 9-12). Зона выделяется в пределах толщ 5-6 свиты мыса Плоского толщ 7-8 юнюньваямской свиты. Нижняя граница устанавливается по последнему нахождению вида индекса предыдущей зоны. Характерны, иногда многочисленные, Cyclammina praecancellata. Зональный комплекс беден по видовому составу. В это время активно проявлялась вулканическая деятельность, водные массы были холодноводными, недосыщенными кальцием и кислородом.
Возможно, как и предыдущая, зона Cyclammina praecancellata соответствует части зоны Islandiella excavata-Haplophragmoides impressus Юго-Западной Камчатки (Дмитриева, Фрегатова, 2004; Дмитриева, 2005). Сопоставляется с нижненутовским подгоризонтом Сахалина, а также с верхней частью этолонского горизонта Западной Камчатки. Зона соответствует верхней части горизонта мыса Плоского и нижней части юнюньваямского горизонта Восточной Камчатки. Верхний миоцен.
VI. Зона Cribroelphidium vulgare (слои 13). Выделяется в пределах толщи 9 юнюньвамской свиты. Нижняя граница устанавливается по появлению вида индекса. Содержит обедненный относительно мелководный комплекс с преобладанием секреционных форм Cribroelphidium vulgare, Quinqueloculina sakhalinica. Отложения зоны соответствует этапу начала резкой перестройки бассейна, связанным с его значительным обмелением. Глубина бассейна составляла до 50-100 м.
Соответствует зоне Islandiella laticamerata Юго-Западной Камчатки (Дмитриева, Фрегатова, 2004; Дмитриева, 2005), зоне Cribroelphidium yabei Северо-Западной Японии (Hanagato, 2003). Комплекс зоны коррелируются с комплексами этолонского горизонта Западной Камчатки, нижненутовского подгоризонта Северного Сахалина, хузинской свиты Пограничного прогиба и маруямского горизонта Юго-Восточного Сахалина. Соответствует верхней части юнюньваямского горизонта. Верхний миоцен.
VII. Зона Elphidiella oregonensis (слои 14, 15). Выделяется в пределах толщи 10 лимимтэвамской свиты. Нижняя граница устанавливается по появлению вида индекса. Характерно преобладание широко распространенных в нижнем плиоцене всей Северной Пацифики Elphidiella oregonensis. Отложения зоны накапливались в условиях новой трансгрессии. Глубины бассейна были около 50 м. Температурные условия были более теплыми, чем в предыдущем этапе.
Соответствует зоне Elphidiella simplex Юго-Западной Камчатки (Дмитриева, Фрегатова, 2004; Дмитриева, 2005), зоне Elphidiella hannai нижнего плиоцена Северо-Восточной Пацифики (Narayan et al, 2005), зоне Elphidium - Cassidulina Северо-Восточной Японии (Hasegawa, 1979). Хорошо сопоставляются с верхненутовским подгоризонтом Северного Сахалина, верхнемаруямским подгоризонтом Южного Сахалина и энемтенским горизонтом Западной Камчатки. Соответствует нижней части лимимтэваямского горизонта. Нижний плиоцен.
VIII. Зона Retroelphidium clavatum (слои 16-21). Выделяется в пределах толщ 11-12 лимимтэваямской свиты, толщи 13 усть-лимимтэваямской свиты и толщи 14 тусатуваямских слоев. Нижняя граница устанавливается по появлению вида индекса. Для комплекса характерно значительное количество Retroelphidium clavatum и Buccella pseudofrigida. Появились Islandiella laticamerata, Bolivina spissa, B. pseudopunctata, Nonionella globosa, Epistominella pacifica. Изменение состава комплекса фораминифер данной зоны связано с последующим углублением бассейна в конце плиоцена. Глубины составляли 100-200 м. Водные массы были насыщены кислородом и кальцием.
Коррелируется с помырским горизонтом Северного Сахалина. Зона соответствует верхней части лимимтэваямского, усть-лимимтэваямскому и тусатуваямскому горизонтам. Верхний плиоцен.
Зона Retroelhidium clavatum была выделена также в ольховской свите п-ва Камчатского (слои 1-4). Характерно значительное количество представителей исландиеллид и эльфидиид. Наряду с бореальными формами, важное место в комплексе занимают аркто-бореальные фораминиферы. Преобладают Retroelohidium clavatum, R. hughesi, Cribroelphidium bartletti. Характеризуется также широким развитием планктонных форм Neogloboquadrina pachydermа, Globigerina. bulloides, G. quinqueloba в средней части зоны. Отложения этой зоны, с учетом данных по диатововым водорослям, соответствует верхней части одноименной зоны о. Карагинского.
|