6.2.1. Микроорганизмы, распространенные в сульфидных рудах
В такой биокосной системе, как окисленные руды, бедной органическим веществом, широко распространены простейшие одноклеточные микроорганизмы (протисты), обычно представленные ветвью низших протистов или прокаристов. К ним относят так называемые хемолитотрофные бактерии, использующие в своих клеточных процессах энергию преобразования (окисления или восстановления) неорганического минерального вещества.
В зависимости от источников углерода, необходимого для воспроизводства биомассы (конструктивный метаболизм), хемолитотрофы делятся на автотрофы, использующие неорганический углерод, и гетеротрофы, "питающиеся" органикой. На рудных месторождениях шире распространены автотрофы, особо характерные для верхних горизонтов профилей окисленных руд.
Месторождениям сульфидных и серных руд более свойственны бактерии семейства серных или тионовых бактерий, окисляющих серу и соединения серы (сульфиды) и сульфатредуцирующие прокариоты. Их классификация и общая характеристика представлены в табл. 46. Перед кратким обзором табличных данных следует представить некоторые самые общие сведения о бактериях, их строении, способе размножения и условиях жизнедеятельности.
Бактерии принадлежат к одноклеточным микроорганизмам, чаще имеющим форму палочки. В меньшей степени распространены клетки шаровой формы (кокки), нередко собранные в цепи (стрептококки) или в скопления изометрической формы (стафилококки). Некоторые виды бактерий известны в виде запятых или завитков (спириллы и вибрионы).
Размер бактериальных клеток составляет доли микрона или микроны. Вес клетки около 4.10-13г, плотность - 1,055г/см3. Химический состав бактерий сложный. Главным компонентом бактериальной клетки является вода (70-85%). В сухом остатке (30-15%) содержатся
Таблица 46
Характеристика главнейших бактерий, распространенных на сульфидных и серных месторождениях
|
Серные (тионовые) и cульфат- редуцирующие бактерии |
Форма, размер (мкм) клетки |
Интервал pH среды |
Окисляемый или восстанавли-ваемый субс-трат |
Тип месторождений |
|
Thiobacillus ferrooxidans
Thiobacillus thiooxidans
Thiobacillus thioparus
Thiobacillus neapolitanus
Thiobacillus "у"
Thiobacillus organoparus
Stibiobacter senarmontii
Сульфитредуци-рующие бакте-рии |
Палочка с одним жгутиком, 0,4х(1-1,5)
Палочка со спи-ралевидным жгу-тиком (0,5-0,8)х1
Палочка с закруг-ленными концами с полярным жгути-ком. Длина до 1,5
Палочка с одним жгутиком, 0,5х1
Палочка с одним жгутиком, 0,5х1
Палочка с закруг-ленными концами с 1-2 субтерми-нальными жгути-ками, (0,5-0,8)х(1-1,5)
Палочка с субтер-минальным жгути-ком 0,5х(0,5-1,8)
Вибрионы, палочки |
1-5
1-5
3-10
3-7
5-9
2-5
5-5,8
4-10 |
S0, SO32-, S2-, S22-, Fe2+
S0, SO32-
S0, H2S
S0, H2S, S22-, Fe2+
S0, S2-
S0, органи-ческие веще-ства
Sb3+
S+6, органика |
Сульфидные и серные руды
Преимущественно серные месторож-дения
То же
Преимущественно пиритизирован-ные угольные
Галенит-антимо-нитовые и висму-тиновые руды
Серные руды
Антимонитовые руды
Сульфидные и серные руды |
органогенные (углерод-50%, кислород-30%, азот до-12%, водород-до
8%) и зольные (натрий, калий, кальций, фосфор, магний, железо и пр. - всего
около 10%) элементы, входящие в состав клеткообразующих органических (белки,
углеводы, жиры и кислоты) и неорганических (фосфаты, нитраты, сульфаты и др.)
соединений. Основа клетки - белки, состоящие из аминокислот. Углеводы и жиры
(липиды) служат источником энергии микроорганизма. Их вид и состав зависит
не только от типа бактерий, но и роста и условий их развития. Среди органических
кислот клетки следует отметить нуклеиновые - дезоксирибонуклеиновую (ДНК)
и рибонуклеиновую (РНК) кислоты, играющие первостепенную роль в наследственной
информации клетки. Фосфатные соединения, производные адениловой кислоты -
аденозинтрифосфат (АТФ), аденозиндифосфат (АДФ), аденозинмонофосфат (АМФ)
и др. принадлежат к числу своеобразных запасников (аккумуляторов) энергии,
служат высокими акцепторами электронов. Механизм их генерации связан с процессами
окисления неорганических веществ, стимулируемый бактериальной клеткой.
Снаружи клетка Thiobacillus имеет жгутики (1-2), размещенные
вдоль или поперек удлинения клетки, и реснички. Вместе они образуют аппарат
движения клетки. Поверхность клетки покрыта слизистой капсулой, служащей средством
захвата и прикрепления к другим особям (образование колоний) и к косному субстрату.
Слизистый слой состоит из воды (98%) и органического (полисахаридного) вещества.
В слизистую капсулу происходит захват минеральных частиц. В ней, начиная со
смачивания минерала слизью капсулы, начинается процесс его окисления. Слизистое
вещество обладает высокой ферментативной активностью.
Форму клетки сохраняет клеточная стенка, являющаяся многослойной
оболочкой высокой прочности, эластичности и упругости. Клеточная стенка состоит
из полимерных белковых и углеводородных комплексов (полипептидов, аминокислот,
полисахаридов). Она защищает клетку от механических воздействий, а также от
проникновения в нее лишней воды и солей. Синтез вещества клеточной стенки
происходит на расположенной под ней еще одной многослойной оболочке клетки
- цитоплазменной мембране, толщина которой всего 50-100Å
. Цитоплазменная мембрана имеет очень сложное строение и выполняет важнейшие
функции, обеспечивающие жизнь клетки. В ней сосредоточены окислительные ферменты,
катализаторы, запасники энергии в виде АТФ, АДФ, АМФ, генерация которых (фосфорилирование)
осуществляется с участием особых веществ (цитохром или гемопротеидов), представляющих
собой соединения кофакторной природы, т.е. содержащие металл переменной валентности,
в данном случае железо. Процесс фосфорилирования, представляющий собой синтез
высокоэнергетических Fe2+-фосфатных связей (АТФ, АДФ и др.), протекает
с участием электронов, переносимых через мембрану с помощью цитохром. Согласно
современным представлениям транспорт электронов через цитоплазменную мембрану
осуществляется в одном направлении, в результате чего возникает довольно высокий
мембранный потенциал клетки. С цитоплазменной мембраной связан энергетический
метаболизм бактерий.
Наконец, основную часть клетки составляет цитоплазма (протоплазма
с диффузным ядром), заключенная в цитоплазменной мембране. В цитоплазме сосредоточен
генетический аппарат клетки: здесь протекают процессы, приводящие клетку к
делению, совершается конструктивный метаболизм, т.е. биохимические превращения,
сопровождаемые накоплением биомассы. Размножение бактерий, происходящее делением,
совершается весьма интенсивно: через 20-30 мин количество клеток обычно удваивается.
За сутки одна особь дает 60-70 поколений.
Тионовые бактерии, распространенные на рудных месторождениях,
принимают участие в окислении соединений с восстановленной формой серы - сульфидов,
арсенидов, серы самородной, а также тиосульфатных и сульфитных образований
в растворе. Большинство из них - аэробы, живущие в кислородсодержащей среде.
Универсальным микроорганизмом, окисляющим сульфиды, является
бактерия Thiobacillus ferrooxidans (рис.8), которая может использовать
в качестве энергетического субстрата практически все сульфидные минералы,
восстановленные соединения серы (So, SO32-
и др.) и другие закисные элементы в растворе, в частности железо. Этот микроорганизм
был открыт около 50 лет назад в кислой шахтной воде, которая после 2-3 дней
стояния на воздухе выделяла ржавый осадок гидроксидов железа. Было доказано,
что окисление сульфата закиси железа, содержащегося в шахтной воде, происходит
с помощью Thiobacillus ferrooxidans.
В настоящее время хорошо изучена морфология и физиология
Th. ferrooxidans. Палочковидные клетки этого организма с одним жгутиком
имеют длину до 1-1,5 мкм при ширине 0,4-0,5 мкм. Размножается этот организм
путем поперечного деления. Для него характерны небольшие колонии, на которых
в растворе обычно возникает янтарно-желтый осадок гидроокислов железа.
Развитие Th. ferrooxidans совершается в сернокислых
растворах с оптимальным значением pH 1,5-3. Окислительный потенциал раствора
при этом достигает 0,75-0,80 В. Установлен прямой контакт клетки Th. ferrooxidans
с минеральным субстратом, частички которого обычно проникают в слизистую капсулу
клетки (Пивоварова, 1977). Источником энергии для этого микроорганизма служат
процессы окисления почти всех сульфидов, самородной серы и восстановленных
форм серы (например, гипосульфита), а также разных закисных металлов в водном
растворе. Определено, что повышенное содержание в растворе органических веществ
(глюкозы и др.) не оказывает задерживающего действия на развитие Th. ferrooxidans.
Также установлено, что клетки этого организма мало проницаемы для некоторых
токсичных металлов, например для меди и цинка, и отличаются сравнительно высокой
адаптацией к среде. Эта культура не спорообразующая.
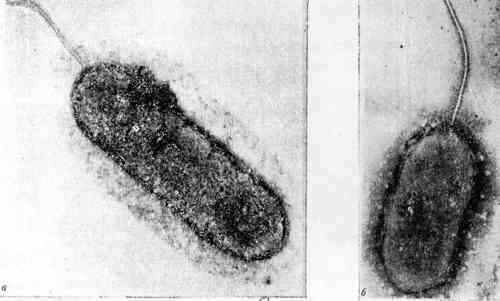 |
Рис. 8. Тиобациллы (электронные
микрофотографии):a - Th. thiooxidans (х90000), b - Th. ferrooxidans
(х90000), (по: Пивоварова, Головачева, 1985) |
В табл. 47 приведены имеющиеся данные по распространенности
Th. ferrooxidans в некоторых типах рудных месторождений. Как видно,
максимальное количество клеток этого микроорганизма приходится на лучше обследованные
медно-колчеданные и колчеданно-полиметаллические руды (в среднем 106-107
кл/г). В полиметаллических месторождениях Сев. Кавказа (Садонская группа)
количество Th. ferrooxidans резко снижено до 10-102
кл/г, что, как уже отмечалось, объясняется малой кислотностью рудничных вод,
нейтрализованных содержащимися в руде карбонатами.
В медно-никелевых месторождениях Норильского и Кольского
районов также установлены очень низкие содержания Th. ferrooxidans
(низкие окислительные потенциалы рудничных вод). Основной причиной служат
холодные климатические условия. Практическое отсутствие Th. ferrooxidans
в золото-арсенопиритовых рудах ряда Казахстанских месторождений (Саяк,
Бакырчик и др.), размещенных в сравнительно благоприятной климатической зоне,
связано с содержанием в рудничных водах токсичного для Th. ferrooxidans
мышьяка (Кулебакин, 1978; и др.).
Два других тионовых микроорганизма - Th. thiooxidans
и Th. thioparus - имеют также форму палочек со спиралевидным
жгутиком. Оба распространены в большей мере на месторождениях серы, которую
Таблица 47
Распространенность микроорганизма Th. ferrooxidans
в рудах некоторых месторождений (данные Н. Н. Ляликовой, С. И. Кузнецова,
Г. И. Каравайко, А. И. Гольбрахта)
|
Месторождения
|
Температу-ра проб, оС
|
pH растворов
|
Eh
|
Кол-во клеток в 1 г руды
|
|
Медно-колчеданные месторождения
|
|
Дегтярское (Ср.Урал)
Блявинское (Ю.Урал)
Кафансоре (Армения)
|
10-21
10-12
15-17
|
2,2-30
0,7-1,7
2,0-2,5
|
0,50-0,78
0,54-0,78
0,75-0,77
|
105-108
106
105
|
|
Колчеданно-полиметаллические месторождения
|
|
Квемо-Болнисское (Грузия)
Маднеульское (Грузия)
Николаевское (Вост.Казахстан)
|
-
-
-
|
3,2-4,5
2,5-3,7
2,5-5,0
|
0,48-0,62
0,52-0,72
-
|
104-107
107
102-105
|
|
Полиметаллические месторождения
|
|
Садонская группа (Сев.Кавказ)
Садон
Худес
Уруп
|
-
-
|
6-7
2-4
3-8
|
-
-
-
|
0-10
102-104
0-103
|
|
Медные вкрапленные руды
|
|
Коунрад (Центр.Казахстан)
|
-
|
3-4
|
-
|
104
|
|
Пиритизированные породы (сланцы, глины)
|
|
Черемшанское и Липовское месторождения
(Ср.Урал)
|
10-15
-
|
3-4
4
|
0,58
0,68
|
105
104
|
|
Медно-никелевые месторождения
|
|
Кольская группа*
Каула
Ниттис-Кумужье
Нюд
Норильская группа
Талнах
|
2-4
2-4
2-4
10-12
|
7-8
6-8
2,4-2,8
6,5-9,0
|
-
0,20-0,40
-
0,20-0,40
|
0-102
0-103
103-104
10-102
|
|
Золото-мышьяковые месторождения
|
|
Саяк, Бакырчик (Казахстан)
|
-
|
-
|
-
|
не обн.
|
*В ряде проб pH=3-4, Eh=0,6-0,8 и число клеток до 107.
окисляют до серной кислоты. Th. thioparus, помимо
серы, окисляет и H2S. Th. thiooxidans развивается в кислых
средах (оптимальная величина pH=2), а Th. thioparus - в нейтрально-щелочных
условиях (pH=7-9) со слабым доступом кислорода. С деятельностью этих тиобацилл
связывается образование серы в осадочных породах при наличии в них подтока
сероводородных (нефтяных) вод. Обследование серных месторождений Гаурдак,
Шор-Су и Раздол показало, что в 1 г руды здесь содержится до 104-105
клеток Th. thiooxidans. Во многих рудных месторождениях, особенно медно-колчеданных
(Урал), медно-молибденовых (Узбекистан) и полиметаллических (Армения), вместе
с Th. ferrooxidans обнаружены высокие концентрации клеток Th. thiooxidans
(до 104-106 кл), участвующих в процессах окисления руд
(Каравайко и др., 1972; и др.). Величины Eh и pH растворов в этих месторождениях
соответственно достигают 0,6 В и 2-3. Культура Th. thioparus в количестве
10-100 кл/мл воды и 10 кл/г руды обнаружена в золото-мышьяковых месторождениях
Казахстана (Гольбрайхт, 1970) и медных рудах Коунрада (Ляликова, Соколова,
1965).
Микроорганизм Thiobacillus "у" развивается
преимущественно в нейтрально-щелочных условиях на галените, висмутине, антимоните
и сере; является строгим автотрофом. Систематические данные по распространенности
в рудах и водах месторождений Pb, Sb и Bi отсутствуют.
Очень мало данных имеется о распространенности на месторождениях
таких тионовых бактерий, как Thiobacillus organoparus и Leptospirillium
ferrooxidans. Th. оrganoparus - короткая, но довольно широкая палочка
с поперечно расположенным жгутиком, способная в кислой среде автотрофно развиваться
на сере и гетеротрофно на легко усваиваемых органических веществах. Второй
отмеченный микроорганизм, представляющий длинную спиралевидную палочку, активно
окисляет железистые сульфиды - пирит и марказит. На рис. 9 показана капля
рудничной воды пиритизированного угольного месторождения, содержащая. Th.
neapolitanus
Открытый Н. В. Ляликовой (1972) микроорганизм Stibiobacter
senarmontii в количестве 103-104 кл/г обнаружен
в сурьмяных месторождениях Ср. Азии (Хайдаркан, Кадамжай и др.) и Украины
(Никитовка). Развиваясь в умеренно-кислой и нейтральной средах, что характерно
для антимонитовых руд, обычно размещенных в карбонатных породах, Stibiobacter
senarmontii окисляет Sb3+ до Sb5+, способствуя тем
самым образованию в окисленных рудах этих месторождений окислов пятивалентной
сурьмы и антимонитов (сервантит, стибиоконит, флайолотит и пр.).
Сульфатредуцирующие бактерии, известные на рудных месторождениях,
объединяются в два главных рода - Desulfotomaculum (спорообразующие) и Desulfovibrio
(неспорообразующие). Развиваются в условиях низкого окислительного потенциала
среды (0,1-0,2 В) в нейтрально-щелочных растворах при наличии органического
вещества (вымывание органики с поверхности, из вмещающих пород, поступление
с нефтяными водами). Известны на больших глубинах (до 3-4 км). По типу дыхания
это анаэробы, использующие углекислый газ и водород. Среди них есть типичные
термофилы, выдерживающие температуру до +90оС. Большинство этих
бактерий устойчиво к высокой концентрации в водах солей (до 300 г/л). Главной
жизнеобеспечивающей функцией служит стимулирование реакций восстановления
сульфатной серы до сероводородной. В результате создаются условия для образования
не только серов |